
Wnt 信号通路
 2019-11-06
2019-11-06 
 MCE小分子
MCE小分子
Wnt 信号通路是一种古老的、进化上保守的通路。其中,Wnt 蛋白属于分泌的糖蛋白,可与卷曲蛋白 (Frizzled) 家族的受体,如与脂蛋白受体相关的蛋白 5/6 (LRP5/6),Ror2 和 Ryk 相互作用,导致各种细胞内信号传导级联激活,这些信号传导级联可以交叉连接或独立发挥作用,调节多种多样的过程,包括细胞增殖,分化,迁移,极性和不对称细胞分裂以及与干细胞更新有关。因此,Wnt 途径的突变往往与人类出生缺陷、癌症以及其他疾病有关。
细胞外 Wnt 信号刺激会引起几种细胞内信号转导级联,包括 Wnt/β-catenin 依赖 (经典) 途径和 β-catenin 独立 (非经典) 途径,后者又可进一步分为平面细胞极性 (PCP) 和 Wnt/Ca2+途径。
β-catenin 依赖 (经典) 信号途径
Wnt/β-catenin 经典途径的标志是粘附连接相关蛋白 β-catenin 向细胞核的积累和易位。在没有 Wnt 信号时,细胞质 β-catenin 被 β-catenin 破坏复合物降解,破坏复合物含有支架蛋白 Axin,APC,GSK3β,CK1。YAP/TAZ 也可以与这种复合物缔合 (对募集 β-TrCP E3 泛素连接酶必不可少)。具体为:CK1 和 GSK-3β 在该复合物中对 β-catenin 磷酸化,从而导致被 β-TrCP E3 连接酶泛素化,并被 26S 蛋白酶体降解。同时,TCF 与 Groucho 的结合抑制了 Wnt 靶基因。
在有 Wnt 信号时 (如 Wnt1, Wnt3a, Wnt8),Wnt 与由 Frizzled 受体和 LRP5/6 组成的受体复合物的结合引发一系列事件,从而靶向破坏 β-catenin 所需的 APC/Axin/GSK3 复合物。具体为:Frizzled 募集的 Dishevelled (Dsh/Dvl) 导致 LRP5/6 磷酸化 (磷酸化由激酶 GSK3 和 CK1γ 介导)。磷酸化的 LRP5/6 将 Axin 募集到膜上,破坏复合物分解,从而导致细胞质中 β-catenin 的稳定和积累。随后,β-catenin 蛋白易位至细胞核,通过取代 TLE/Groucho 复合物并募集组蛋白修饰共激活物 (如 CBP/p300,BRG1,BCL9 和 Pygo) 与 LEF 和 TCF 蛋白形成活性复合物,调节靶基因的表达。
典型的 Wnt 信号的激活可以被一些 Wnt 蛋白抑制剂抑制,如 sFRP,Dkk,WIF,Wise/SOST,Cerberus,Tiki1 等,它们的共同特征是通过阻止配体-受体相互作用或 Wnt 受体成熟来拮抗 Wnt 信号传导。而 Wnt 信号通路也会被 R-spondin 家族蛋白和 Norrin 蛋白激活。另外,其下游途径的激活也可以被抑制,例如 TAK1-NLK 阻断了 β-catenin 诱导的转录活性。Hippo 途径也可通过 TAZ 抑制 Wnt 信号传导,具体为 TAZ 通过抑制 CK1δ/β 介导的 DVl 磷酸化来限制 Wnt 信号传导。
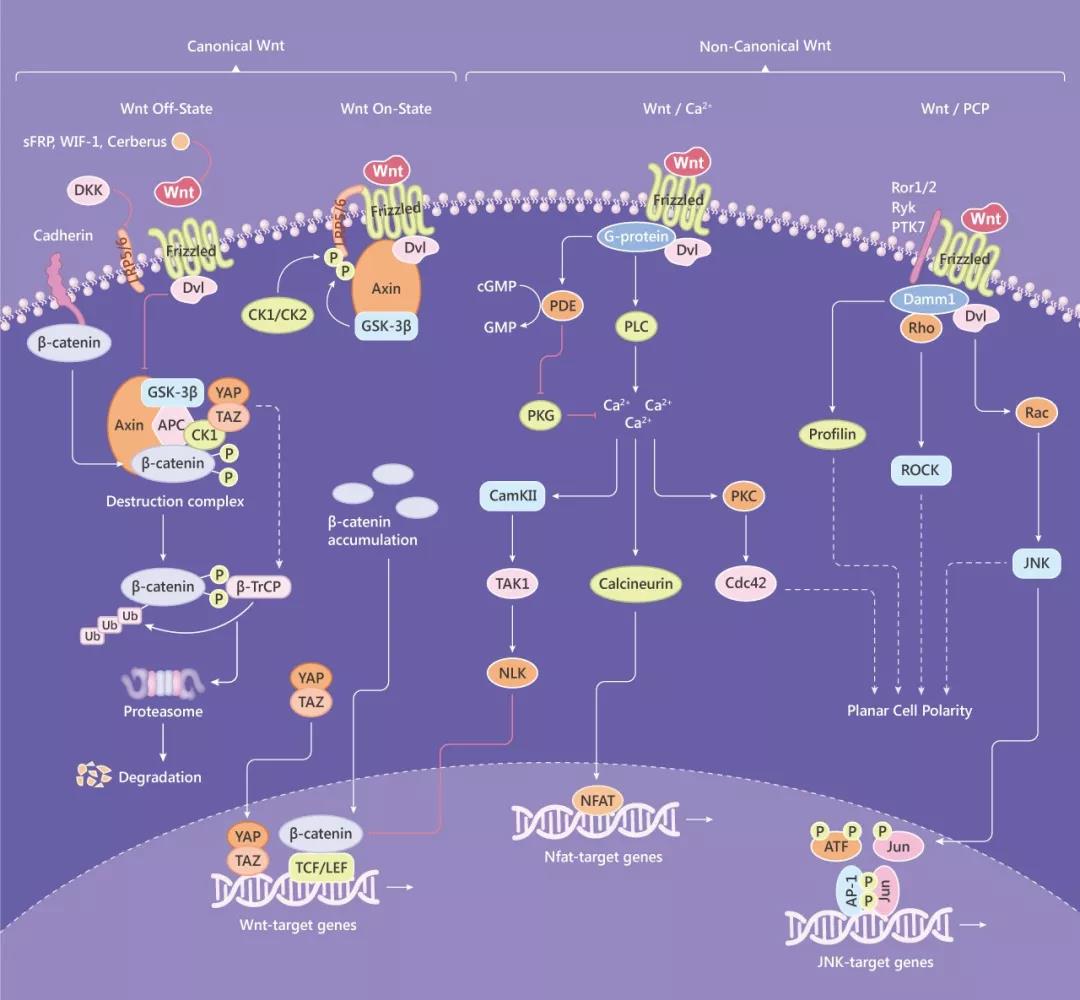
Fig. 1 典型和非典型 Wnt 信号通路图
非经典平面细胞极性 (PCP) 途径
在 PCP 信号传导途径中,Wnt 配体 (如 Wnt5a,Wnt11) 与 Frizzled 受体或其共受体(如 ROR-Frizzled) 结合从而引发级联反应。结合后的受体会募集并激活的 Dvl 蛋白,Dvl 通过 Daam1 介导 Rho 的激活,Rho 的激活又激活 Rho 激酶 (ROCK)。Dvl 还介导了Rac 的激活,从而激活了 JNK。这些导致细胞骨架的重排和/或转录反应,如 ATF2。另外,Daam1 还可通过肌动蛋白结合蛋白 Profilin 介导肌动蛋白聚合。PCP 和β-catenin 依赖的 Wnt 信号可以互相拮抗,抑制一个通常会上调另一个。如,优先激活 PCP 信号的 Wnt5a 会竞争并抑制 Wnt3a 与 Frizzled 2 的结合,从而抑制了 β-catenin 依赖性途径。
非经典 Wnt/Ca2+途径
该途径是由 Frizzled 介导的,通过 G 蛋白激活来激活 Dvl。Dvl 激活磷酸二酯酶 PDE,后者抑制 PKG (阻断未刺激细胞的 Ca2+ 释放),从而使 Ca2+ 水平增加。Dvl 还通过 PLC 激活 IP3,IP3 触发 Ca2+ 从细胞内释放并激活触发细胞内储存的 Ca2+ 释放,也会导致 Ca2+ 水平增加。诱导的钙离子通量可以激活第二信使,如蛋白激酶 C (PKC),钙调蛋白依赖性激酶 II (CamKII) 或钙依赖性磷酸酶钙调神经磷酸酶 (Calcineurin, CaN)。另外,CamKII 激活的 TAK-NLK 可通过 TCF 拮抗 Wnt/β-catenin 信号传导。而活化的 Calcineurin 可以使活化的 T 细胞核因子 (NFAT) 去磷酸化,促进 NFAT 进入细胞核,激活其靶基因。同时,PKC 成员可以激活小 GTPase Cdc42,后者又可以进入 PCP 途径。
Wnt抑制剂/激动剂
Salinomycin,一种离子载体抗生素,同时也是 Wnt/β-catenin 信号的抑制剂。Salinomycin 作用于 Wnt/Fzd/LRP 复合物,阻断 Wnt 诱导的 LRP6 磷酸化,导致 Wnt 共受体 LRP6 蛋白降解。Salinomycin 具有较强的抗癌活性,可选择性抑制人肿瘤干细胞。
IWP-2 是 Wnt 加工和分泌的抑制剂,靶向膜结合 O- 酰基转移酶 porcupine (Porcn),Porcn 是膜结合的 O-酰基转移酶 (MBOAT) 家族的成员,可在 Wnt 蛋白上添加棕榈酰基,这对于其信号传递能力至关重要,并且是 Wnt 分泌所必需的。IWP-2 阻止 Wnt 配体棕榈糖基化阻断以及 Wnt 蛋白的分泌,从而抑制下游 Wnt 信号传导。
XAV939 是一种细胞可穿透性的 Wnt/β-catenin 信号转导抑制剂,通过抑制聚 ADP-核糖基化酶 tankyrase1 和 tankyrase 2 来稳定破坏复合物成分 Axin,从而刺激 β-catenin 降解。
SKL2001 是一种通过增加 β-catenin 的稳定性来激活 Wnt 信号传导的特异的 Wnt/β-catenin 激动剂。SKL2001 与 Axin 竞争性地与 β-catenin 的结合,破坏 Axin/β-catenin 的相互作用,从而破坏 CK1 和 GSK-3β 介导的 Ser33/37/ Thr41 和 Ser45 处 β-catenin 的磷酸化,进而增加 β-catenin 的稳定性,确保其不被蛋白酶体降解。CK1 和 GSK-3β 酶活性不受 SKL2001 影响 (如下图)。
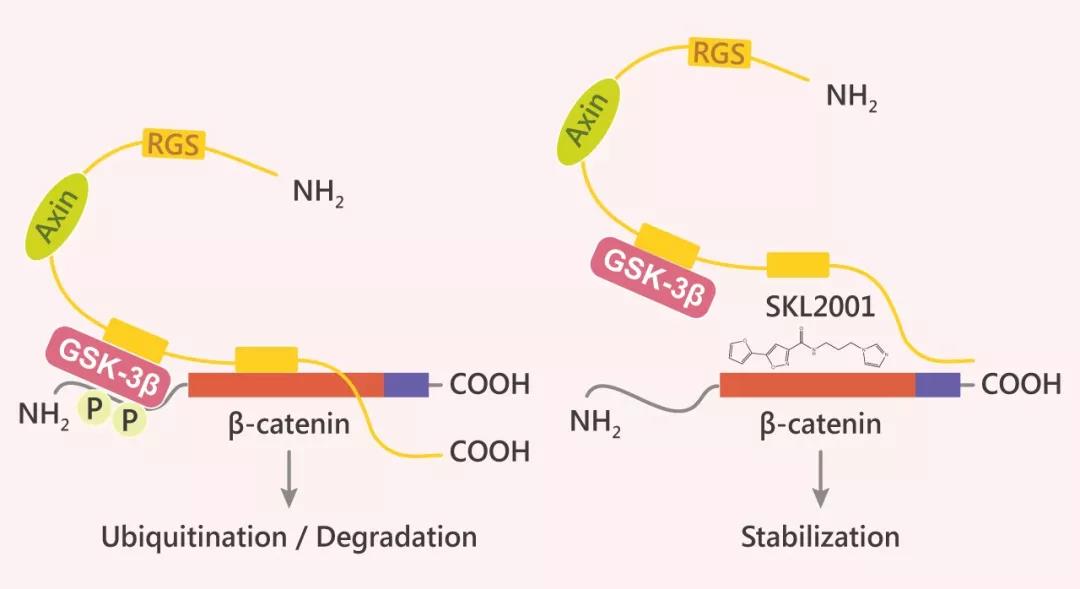
Fig. 2 SKL2001 作用机制图

